

 Printer-friendly page
Printer-friendly page
Volume 5, Issue A ::: August 2001
The Neurobiology of Reading and Dyslexia
by Sally E. Shaywitz, M.D., and Bennett A. Shaywitz, M.D.
Developmental dyslexia is characterized by an unexpected difficulty in reading experienced by children and
adults who otherwise possess the intelligence and motivation considered necessary for
accurate and fluent reading. It represents one of the most common problems
affecting children and adults; in the United
States, the prevalence of dyslexia is estimated to range
from five to 17 percent of school-aged children, with as many as 40
percent of the entire population reading below grade level. Dyslexia (or
specific reading disability) is the most common and most carefully studied of
the learning disabilities, affecting 80 percent of all individuals identified
as learning disabled. This article reviews recent advances in the
neurobiology of dyslexia and their implications for teaching adults with
dyslexia.
Epidemiology of Dyslexia
Like hypertension and obesity, dyslexia fits a dimensional model: within the population, reading and reading disability occur along a continuum, with reading disability representing the lower tail of a normal distribution of reading ability. Good evidence based on sample surveys of randomly selected populations of children now indicate that dyslexia affects boys and girls equally (Figure 1); the long-held belief that only boys suffer from dyslexia reflected sampling bias in school-identified samples.
Dyslexia is a persistent, chronic condition; it does not represent a transient "developmental lag" (Figure 2). Over time, poor readers and good readers tend to maintain their relative positions along the spectrum of reading ability.
Causes
Dyslexia is both familial and heritable: both environmental and genetic influences affect the expression of dyslexia. This observation provides opportunities for early identification of affected siblings and often for delayed but helpful identification of affected adults. Thus 23 to 65 percent of children who have a parent with dyslexia, 40 percent of siblings of dyslexics, and 27 to 49 percent of parents of dyslexics may have the disorder. Studies implicate loci on chromosomes 6 and 15 and, more recently, on chromosome 2 in the causation of dyslexia.
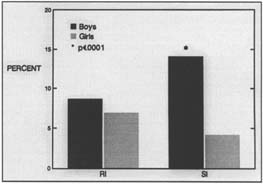
Figure 1. Prevalence of reading disability in research-identified (RI) and school-identified (SI) boys and girls. Schools identify about four times as many boys as girls, reflecting primarily externalizing behavioral characteristics that are more likely to bring boys to a teacher's attention. This skewed prevalence rate reflects referral bias. When actual reading scores are used to identify children, there is no significant difference in the prevalence of dyslexia between boys and girls (based on data in Shaywitz et al., 1990).
The Cognitive Basis of Dyslexia
The phonologic deficit hypothesis - There is now a strong consensus among investigators in the field that the central difficulty in dyslexia reflects a deficit within the language system, although other systems and processes may also contribute to the difficulty. The language system is conceptualized as a hierarchical series of components: at higher levels are neural systems engaged in processing, for example, semantics, syntax, and discourse; at the lowest level is the phonologic module dedicated to processing the distinctive sound elements that constitute language. The functional unit of the phonologic module is the phoneme, defined as the smallest discernible segment of speech; for example, the word "bat" consists of three phonemes: /b/ /ae/ /t/ (buh, aah, tuh). To speak a word, the speaker retrieves the word's phonemic constituents from his or her internal lexicon, assembles the phonemes, and then utters the word. Conversely, to read a word, the reader must first segment that word into its underlying phonologic elements. The awareness that all words can be decomposed into these basic elements of language (phonemes) allows the reader to decipher the reading code. In order to read, a child has to develop the insight that spoken words can be pulled apart into phonemes and that the letters in a written word represent these sounds. This so-called phonemic awareness is largely missing in dyslexic children and adults. Results from large and well-studied populations with reading disability confirm that in young school-aged children, as well as in adolescents, a deficit in phonology represents the most robust and specific correlate of reading disability. Such findings form the basis for the most successful and evidence-based interventions designed to improve reading. While children and adults with a phonologic deficit represent the vast majority of subjects with dyslexia, other subtypes may account for some cases of dyslexia. Examples include dyslexia resulting from deficits in naming-speed in addition to phonological deficits, the so called double-deficit hypothesis.
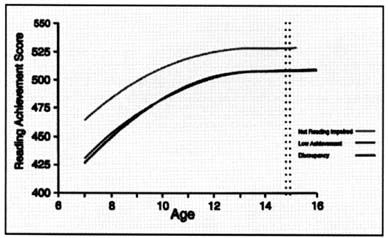
Figure 2. Trajectory of reading skills over time in nonimpaired and dyslexic readers. Ordinate shows Rasch scores (W scores) from the Woodcock-Johnson reading test (Woodcock & Johnson, 1989) and abscissa shows age in years. Both dyslexic and nonimpaired readers improve their reading scores as they get older, but the gap between the dyslexic and nonimpaired readers remains. Thus dyslexia is a deficit and not a developmental lag (from Francis et al., 1996).
Implications of the phonologic model of dyslexia - Reading is comprised of two main processes: decoding and comprehension. In dyslexia, a deficit at the level of the phonologic module impairs the reader's ability to segment the written word into its underlying phonologic elements. As a result, the reader experiences difficulty, first in decoding the word and then in identifying it. The phonologic deficit is domain-specific; that is, it is independent of other, nonphonologic, abilities. In particular, the higher-order cognitive and linguistic functions involved in comprehension, such as general intelligence and reasoning, vocabulary, and syntax, are generally intact. This pattern - a deficit in phonologic analysis contrasted with intact higher-order cognitive abilities - offers an explanation for the paradox of otherwise intelligent people who experience great difficulty in reading.
According to the model, a circumscribed deficit in a lower-order linguistic (phonologic) function blocks access to higher-order processes and to the ability to draw meaning from text. The dyslexic reader cannot use his or her higher-order linguistic skills to access the meaning until the printed word has first been decoded and identified. For example, readers who know the precise meaning of the spoken word "apparition" will not be able to use their knowledge of the meaning of the word until they can decode and identify the printed word on the page and will appear not to know the word's meaning.
The phonologic deficit in adolescence and adult life - Deficits in phonological coding continue to characterize dyslexic readers even in adolescence; performance on phonological processing measures contributes most to differentiating dyslexic from average readers, and average from superior readers as well. Children with dyslexia neither spontaneously remit nor do they demonstrate a lag mechanism for "catching up" in the development of reading skills. That is not to say that many dyslexic readers do not become quite proficient in reading a finite domain of words in their area of special interest, usually words that are important for their careers. Such individuals, while able to decode words in this domain, still exhibit evidence of their early reading problems when they have to read unfamiliar words, which they do accurately but not fluently and automatically. In adolescents, oral reading, the rate of reading, as well as facility with spelling may be most useful clinically in differentiating average from poor readers.
From a clinical perspective, these data indicate that as children approach adolescence, a manifestation of dyslexia may be a very slow reading rate. Children may learn to read words accurately, but they will not be fluent or automatic, reflecting the lingering effects of a phonologic deficit. Because they are able to read words accurately (albeit very slowly), dyslexic adolescents and young adults may mistakenly be assumed to have "outgrown" their dyslexia. These older dyslexic students may be similar to their unimpaired peers on untimed measures of word recognition, yet continue to suffer from the phonologic deficit that makes reading less automatic, more effortful, and slow. The provision of extra time is therefore an essential accommodation; it allows them the time to decode each word and to apply their unimpaired higher-order cognitive and linguistic skills to the surrounding context to get at the meaning of words that they cannot entirely or rapidly decode.
Neurobiological Influences
A range of neurobiological investigations using postmortem brain specimens and, more recently, brain morphometry and diffusion tensor magnetic resonance imaging (MRI) suggests that there are differences between dyslexic and nonimpaired readers in the back of the brain, specifically in the temporoparieto-occipital brain regions. Functional brain imaging studies also show a failure of left hemisphere posterior brain systems to function properly in adult dyslexic readers while they perform reading tasks.
In principle, functional brain imaging is quite simple. When an individual is asked to perform a discrete cognitive task, that task places processing demands on particular neural systems in the brain. To meet those demands requires activation of neural systems in specific brain regions and those changes in neural activity are, in turn, reflected by changes in cerebral blood flow. We use the term "functional imaging" for technologies that measure those changes in blood flow in specific brain regions while subjects are engaged in cognitive tasks.
Gender-Based Differences
In an early study of 19 neurologically normal right‑handed men and 19 women, the subjects had to decide whether two pseudowords rhymed. (For example, do [LEAT] and [JETE] rhyme?) Nonword reading is perhaps the clearest indication of decoding ability because familiarity with the letter pattern cannot influence the individual's response. Of particular interest were differences in brain activation patterns in men compared to women. Figure 3 illustrates that activation during phonological processing in men was more lateralized to the left inferior frontal gyrus, known as Broca's area; in contrast, activation during this same task in women resulted in a more bilateral pattern of activation of this region.
These findings provide the first clear evidence of gender-based differences in the functional organization of the brain for language. They support and extend a long-held hypothesis that language functions are more likely to be highly lateralized in males but are represented in both cerebral hemispheres in females.
Studies of dyslexic readers indicate a significant disruption in the neural systems for reading in dyslexic subjects as they try to decode pseudowords. Thus, as shown in Figure 4 during nonword rhyming in dyslexic readers, we found a disruption in several critical components of a posterior system involving the posterior superior temporal gyrus (Wernicke's area) and the angular gyrus, and a concomitant increase in activation in the inferior frontal gyrus.
These data indicate that dyslexic readers demonstrate a functional disruption in an extensive system in the posterior cortex encompassing both traditional visual and language regions as well as a portion of association cortex. The involvement of this latter region, centered about the angular gyrus, is of particular interest since this portion of association cortex is considered pivotal in carrying out those cross-modal integrations necessary for reading (i.e., mapping the visual percept of the print onto the phonologic structures of the language).
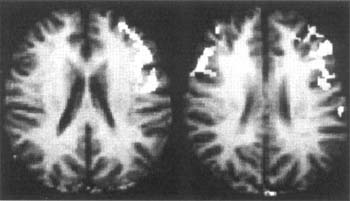
Figure 3. Gender-based differences in the brain during phonological processing. Composite fMRI images show the distribution of brain activation patterns in men (left) and women (right) during a nonword rhyming task. In men, activation is lateralized to the left inferior frontal regions; in women the same region is active bilaterally (data from Shaywitz et al., 1995).
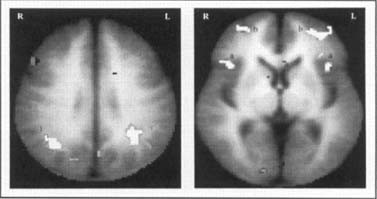
Figure 4. Composite fMRI activation maps in nonimpaired and dyslexic readers engaged in phonological processing during the nonword rhyme task show that nonimpaired readers activate a large region involving the angular gyrus (1), supramarginal gyrus, and posterior portions of the superior temporal gyrus. In contrast, dyslexic readers demonstrate a relative underactivation in this posterior region and an increased activation in the inferior gyrus (a) and middle front gyrus (b) bilaterally (data from Shaywitz et al., 1998).
Consistent with this study of developmental dyslexia, a large literature on acquired inability to read (alexia, for example, following a stroke) describes neuroanatomical lesions most prominently centered about the angular gyrus. It should not be surprising that both the acquired and the developmental disorders affecting reading have in common a disruption within the neural systems serving to link the visual representations of the letters to the phonologic (language) structures they represent. While reading difficulty is the primary symptom in both acquired alexia and developmental dyslexia, associated symptoms and findings in the two disorders would be expected to differ somewhat, reflecting the differences between an acquired and a developmental disorder. In acquired alexia, a structural lesion resulting from an insult (e.g., stroke, tumor) disrupts a component of an already functioning neural system and the lesion may extend to involve other brain regions and systems. In developmental dyslexia, as a result of a constitutionally based functional disruption, the system never develops normally. The symptoms reflect the emanative effects of an early disruption to the phonologic system. In either case the disruption is within the same neuroanatomical system.
A Neural Model for Reading
These data from laboratories around the world indicate that a number of interrelated neural systems are used in reading: at least two in posterior brain regions as well as distinct and related systems in anterior regions (Figure 5).
In order to read, the beginning reader must break the reading code, that is, transform the visual features (the letters) of the word into the linguistic sounds (the phonemes) they represent and then access the meaning of the word. As early as 1891, Dejerine suggested that a portion of the posterior brain region (which includes the angular gyrus and supramarginal gyrus in the inferior parietal lobule, and the posterior aspect of the superior temporal gyrus) is critical for reading.
Rather than the smoothly functioning and integrated reading systems observed in nonimpaired readers, disruption of the posterior reading systems results in dyslexic readers attempting to compensate by shifting to other, ancillary, systems (e.g., anterior sites such as the inferior frontal gyrus and right posterior sites). The anterior sites, which are critical in articulation, may help dyslexic readers develop an awareness of the sound structure of the word by forming the word with their lips, tongue, and vocal apparatus and thus allow them to read, albeit more slowly and less efficiently than if the fast occipitotemporal word identification system were functioning. The posterior sites, for example the right occipitotemporal area, may be used by the dyslexic reader to facilitate visual pattern recognition, compensating for the impaired word analysis systems in the left posterior regions. The shift to ancillary neural systems in dyslexic readers may support accurate, but not fluent and automatic, word reading.
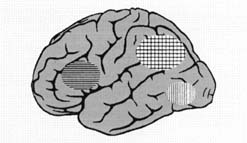
Figure 5. Neural systems for reading. Converging evidence indicates three important systems in reading, all primarily in the left hemisphere: 1) anterior system in the left inferior frontal region; 2) dorsal parietotemporal system involving angular gyrus, supramarginal gyrus, and posterior portions of the superior temporal gyrus; 3) ventral occipitotemporal system involving portions of the middle temporal gyrus and middle occipital gyrus. See text for details.
Delineation of the circuitry for reading in dyslexia may now allow strategies for specific interventions designed to facilitate the function of these ancillary systems, and a method to measure the efficacy of such interventions in a more focused and efficient way. Such studies are now underway.
For dyslexic readers, these brain activation patterns provide evidence of an imperfectly functioning system for segmenting words into their phonologic constituents; accordingly, this disruption is evident when dyslexic readers are asked to respond to increasing demands on their phonologic analysis. These findings now add neurobiological support for previous cognitive/behavioral data, pointing to the critical role of phonologic analysis, and its impairment, in dyslexia. The pattern of relative underactivation in posterior brain regions contrasted with relative overactivation in anterior regions may provide a neural signature for the phonologic difficulties characterizing dyslexia.
Editor's note: Portions of this chapter appeared in (Shaywitz 1998; Shaywitz and Shaywitz 1999; Shaywitz, Pugh et al. 2000; Shaywitz, Shaywitz et al. In Press; Shaywitz, Shaywitz et al. In Press; Shaywitz, Lyon et al. In Press) with permission.
Acknowledgments
This
work was supported by grants from the National Institute
of Child Health and Human Development (PO1 HD 21888 and P50 HD25802). We thank
Carmel Lepore for her help in preparing the manuscript.
References
Anderson, A., & Gore, J. (1997). "The physical basis of neuroimaging techniques." In M. Lewis and B. Peterson (eds.), Child and Adolescent Psychiatric Clinics of North America (vol. 6, pp. 213-264). Philadelphia: W. B. Saunders Co.
Bruck, M. (1992). "Persistence of dyslexics' phonological awareness deficits." Developmental Psychology, 28(5), 874-886.
Filipek, P. (1996). "Structural variations in measures in the developmental disorders." In R. Thatcher, G. Lyon, J. Rumsey, and N. Krasnegor (eds.), Developmental Neuroimaging: Mapping the Development of Brain and Behavior (pp. 169-186). San Diego, CA: Academic Press.
Frackowiak, R., Friston, K., et al. (1997). Human Brain Function. New York: Academic Press.
Francis, D. J., Shaywitz, S. E., et al. (1996). "Developmental lag versus deficit models of reading disability: A longitudinal, individual growth curves analysis." Journal of Educational Psychology, 88(1), 3-17.
Panel, R. o. t. N. R. (2000). Teaching Children to Read: An Evidence-Based Assessment of the Scientific Research Literature on Reading and its Implications for Reading Instruction. Bethesda, MD: National Institute of Child Health and Human Development, National Institutes of Health.
Paulesu, E., Demonet, J. F., et al. (2001). "Dyslexia - cultural diversity and biological unity." Science, 291, 2165-2167.
Pennington, B. F., and Gilger, J. W. (1996). "How is dyslexia transmitted?" In C. H. Chase, G. D. Rosen, and G. F. Sherman (eds.), Developmental Dyslexia. Neural, Cognitive, and Genetic Mechanisms (pp. 41-61). Baltimore: York Press.
Shaywitz, B., Pugh, K. R., et al. (2000). "The neurobiology of reading and reading disability (dyslexia)." In M. Kamil, P. Mosenthal, P. Pearson, and R. Barr (eds.), Handbook of Reading Research (vol. III, pp. 229-249). Mahwah, NJ: Lawrence Erlbaum Associates.
Shaywitz, B., Shaywitz, S., et al. (1995). "Sex differences in the functional organization of the brain for language." Nature, 373, 607-609.
Shaywitz, S. (1998). "Current concepts: Dyslexia." The New England Journal of Medicine, 338(5), 307-312.
Shaywitz, S., & Shaywitz, B. (1999). "Dyslexia". In K. Swaiman and S. Ashwal (eds.), Pediatric Neurology: Principles & Practice (vol. 1, pp. 576-584). St. Louis, MO: Mosby.
Shaywitz, S., Shaywitz, B., et al. (In Press). "The neurobiology of dyslexia." Clinical Neuroscience Research.
Shaywitz, S., Shaywitz, B. et al. (In Press). "The neuropsychology of dyslexia." In S. Segalowitz and I. Rapin (eds.), Handbook of Neuropsycholology. Amsterdam: Elsevier.
Shaywitz, S.E. (1996). "Dyslexia." Scientific American, 275(5), 98-104.
Shaywitz, S., Fletcher, E., et al. (1999). "Persistence of dyslexia: The Connecticut Longitudinal Study at adolescence." Pediatrics, 104, 1351-1359.
Shaywitz, S., Lyon, E., et al. (In Press). "Dyslexia (specific reading disability)." In F. Burg, J. Ingelfinger, R. Polin, and A. Gershon (eds.), Current Pediatric Therapy. Philadelphia: W.B. Saunders.
Shaywitz, S., Shaywitz, B., et al. (1990). "Prevalence of reading disability in boys and girls: Results of the Connecticut Longitudinal Study." Journal of the American Medical Association, 264(8), 998-1002.
Shaywitz, S. E., Shaywitz, B. A., et al. (1998). "Functional disruption in the organization of the brain for reading in dyslexia." Proceedings of the National Academy of Science of the United States of America, 95, 2636-2641.
Snowling, M. (2000). Dyslexia. Oxford, UK: Blackwell Publishers, Inc.
Torgesen, J. K. (1995). Phonological Awareness: A Critical Factor in Dyslexia, Orton Dyslexia Society.
Wagner, R., and Torgesen, J. (1987). "The nature of phonological processes and its causal role in the acquisition of reading skills." Psychological Bulletin, 101, 192-212.
Woodcock,
R.W., and Johnson, M.B.
(1989). Woodcock-Johnson Psycho-Educational Battery - Revised (WJ-R). Allen,
TX: Developmental Learning Materials.
About the Authors
Sally
E. Shaywitz, M.D., is a Professor of Pediatrics and the Co-Director of the NICHD-Yale
Center for the Study of Learning and Attention.
Bennett
A. Shaywitz, M.D., is a Professor of Pediatrics and Neurology and the
Co-Director of the NICHD-Yale Center for the Study of Learning and Attention.
